Dele:



Hva er polygen arv? (med eksempler)
den polygen arv er overføring av tegn hvis manifestasjon avhenger av flere gener. Ved monogen arv manifesterer seg et tegn fra uttrykket av et enkelt gen; i Digénica, to. I polygen arv snakker vi vanligvis om deltakelse av to, om ikke tre eller flere gener.
Faktisk er det svært få tegn som avhenger av manifestasjonen av bare ett gen eller to gener. Enkelheten i analysen av tegn som er avhengig av noen få gener, har imidlertid hjulpet Mendels arbeid sterkt.

Etterfølgende studier av andre forskere viste at biologisk arv generelt sett er litt mer kompleks enn det.
Når vi snakker om arv av et tegn som avhenger av flere gener, sier vi at de samhandler med hverandre for å gi slike karakterer. I disse interaksjonene komplementerer eller genererer disse generene.
Ett gen kan utføre en del av arbeidet, mens andre utfører en annen. Setningen av handlinger er endelig observert i karakteren av hvis manifestasjon de deltar.
I andre arv bidrar hvert gen med lignende funksjon litt hver til karakterens endelige manifestasjon. I denne klassen av polygen arv blir en additiv effekt alltid observert. I tillegg er variasjonen i tegnets manifestasjon kontinuerlig, ikke diskret.
Til slutt bestemmer fraværet av ekspresjon av et supplementært gen ikke nødvendigvis et fenotype-tap på grunn av fravær, mangel eller nullitet.
index
- 1 Eksempler på polygena tegn
- 1.1 høyde
- 1.2 Animal pels
- 1.3 Sykdommer
- 2 komplementære gener
- 2.1 Epistatiske interaksjoner
- 2.2 Ikke-epistatiske interaksjoner mellom komplementære gener
- 3 Supplerende gener
- 3.1 Noen eksempler på supplerende gener
- 4 referanser
Eksempler på polygena tegn
I de enkleste manifestasjonene er fenotypen alt eller ingenting. Det vil si at det presenterer eller ikke er slik aktivitet, funksjon eller karakteristikk. I andre tilfeller er det to alternativer: for eksempel grønn eller gul.
høyde
Men det er andre tegn som manifesterer seg på en bredere måte. For eksempel, staturen. Tydeligvis har vi alle en statur. Avhengig av det, er vi klassifisert på en bestemt måte: høy eller lav.
Men hvis vi analyserer en befolkningsbrønn, innser vi at det er et svært bredt spekter av høyder - med ekstremer på begge sider av en normal fordeling. Høyde avhenger av manifestasjonen av mange forskjellige gener.
Det avhenger også av andre faktorer, og det er derfor høyden er et tilfelle av polygen og multifaktorial arv. Så mange gener kan måles og involveres, de kraftige verktøyene for kvantitativ genetikk brukes til analyse. Spesielt i analysen av kvantitativt trekksted (QTL, for sin akronym på engelsk).
Pels av dyr
Andre tegn som generelt er polygener, inkluderer manifestasjon av pelsfarge i noen dyr, eller formen av frukten i planter.
Generelt, for enhver karakter hvis manifestasjon viser en rekke kontinuerlig variasjon i befolkningen, kan polygen arv bli mistenkt.
sykdommer
I medisin er studien av genetiske grunnlaget for sykdommer veldig viktig for å forstå dem og finne måter å lindre dem på. I polygen epidemiologi prøver vi for eksempel å bestemme hvor mange forskjellige gener som bidrar til manifestasjon av en sykdom.
Fra dette kan strategier foreslås for å oppdage hvert gen, eller å behandle mangelen på en eller flere av dem.
Enkelte sykdommer i polygenic arv hos mennesker inkluderer astma, schizofreni, noen autoimmune sykdommer, diabetes, hypertensjon, bipolar lidelse, depresjon, hudfarge, etc..
Komplementære gener
Opplevelsen og bevisene som er akkumulert gjennom årene, indikerer at mange gener er involvert i manifestasjon av tegn med flere fenotyper..
I tilfelle komplementære geninteraksjoner mellom alleler av gener fra forskjellige loci, kan disse være epistatisk eller ikke-epistatisk.
Epistatiske interaksjoner
I epistatiske interaksjoner maskerer uttrykket av allelen av et gen fra ett locus uttrykket av et annet fra et annet sted. Det er den vanligste samspillet mellom forskjellige gener som kodes for samme karakter.
For eksempel er det mulig at for et tegn å manifestere, avhenger det av to gener (En/til og B/b). Dette betyr at for å få karakteren til å manifestere, må produktene fra genene delta En og B.
Dette kalles dobbelt dominant epistase. I et tilfelle av recessiv epistase av til på B, Tvert imot, mangelen på manifestasjon av egenskapen kodet av En unngå uttrykk for B. Det er mange forskjellige tilfeller av epistase.
Ikke-epistatiske interaksjoner mellom komplementære gener
Avhengig av hvordan de er definert, er det andre interaksjoner mellom komplementære gener som ikke er epistatisk. Ta for eksempel definisjonen av fjærfarge i fugler.
Det har blitt sett at den biosyntetiske banen som fører til produksjon av et pigment (for eksempel gul), er uavhengig av en annen farge (for eksempel blå).
Både i veien for manifestasjonen av den gule fargen og den blå, som er uavhengige av hverandre, er gen-samspillet epistatisk for hver farge.
Men hvis vi vurderer fargen på fuglens kappe som helhet, er bidraget fra den gule fargen uavhengig av bidraget fra blått. Derfor er manifestasjonen av en farge ikke epistatisk over den andre.
I tillegg er det andre gener som bestemmer mønsteret hvor farger på hud, hår og fjær vises (eller ikke vises). Fargetegnene og fargemønsteret kompletterer hverandre i fargen som den enkelte viser.
På den annen side involverte i hudfarge i mennesker minst tolv forskjellige gener. Det er lett å forstå da hvordan mennesker varierer så mye i farge hvis vi i tillegg legger til andre ikke-genetiske faktorer. For eksempel, sol eksponering (eller kunstige kilder til "soling"), tilgjengelighet av vitamin D, etc..
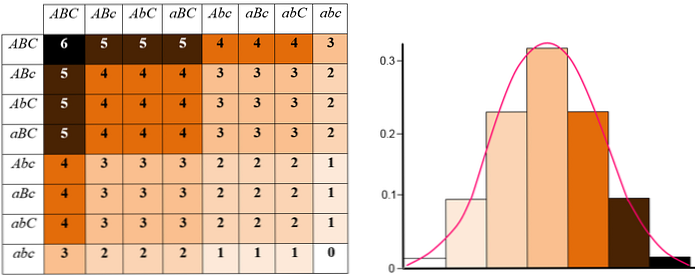
Supplerende gener
Det er tilfeller der et gens handling gjør det mulig å observere manifestasjon av et tegn i større grad. Det er også mulig at det ikke er noe gen å definere en biologisk egenskap som faktisk er summen av mange uavhengige aktiviteter.
For eksempel høyde, melkproduksjon, frøproduksjon, etc. Mange aktiviteter, funksjoner eller kapasiteter legger til for å gi slike fenotyper.
Disse fenotyper er generelt sagt at partene innser manifestasjon av en helhet som viser resultatet av en enkeltperson, en avstamning, avle et dyr, en plante variasjon, etc..
Virkningen av supplerende gener betyr også at det finnes en rekke fenotyper som nesten alltid er definert av en normal fordeling. Noen ganger er det svært vanskelig å skille eller skille komplementær effekt av det kompletterende genet i komplekse fenotyper.
Noen eksempler på supplerende gener
Det har vist seg at virkningen og reaksjonen på visse stoffer, for eksempel, avhenger av aktiviteten til mange forskjellige gener.
Generelt har disse gener også mange alleler i befolkningen, og derfor øker mangfoldet av responser. En lignende sak oppstår i andre tilfeller der en person får vekt ved å konsumere den samme maten mot hvilken en annen ikke opplever betydelige endringer.
Det bør til slutt tilsettes at i tillegg til additivvirkninger som noen gener presenterer, er det de som undertrykker andres manifestasjon.
I disse tilfellene kan et gen ikke relatert til manifestasjon av en annen føre til inaktivering av den første ved genetiske og epigenetiske interaksjoner.
referanser
- Delmore, K. E., Toews, D. P., Germain, R. R., Owens, G. L., Irwin, D. E. (2016) Genetikken av sesongmessige migrering og fjærdrakt farge. Nåværende biologi, 26: 2167-2173.
- Dudbridge, F. (2016) Polygen epidemiologi. Genetisk epidemiologi, 4: 268-272.
- Quillen, EE, Norton, HL, Parra, EJ, Lona-Durazo, F., Ang, KC, Illiescu, FM, Pearson, LN, Shriver, MD, Lasisi, T., Gokcumen, O., Starr, I., Lin., YL, Martin, AR, Jablonski, N. G. (2018) Nyanser av kompleksitet: Nye perspektiver på evolusjon og genetisk arkitektur av menneskelig hud. Amerikansk Journal of Physical Anthropology, doi: 10.1002 / ajpa.23737.
- Maurer, MJ, Sutardja, L., Pinel, D., Bauer, S., Muehlbauer, AL, Ames, TD, Skerker, JM, Arkin, AP (2017) kvantitativ egenskap loci (QTL) -Guidede metabolsk prosjektering av et kompleks egenskap. ACS Syntetisk Biologi, 6: 566-581.
- Sasaki, A., Ashikari, M., Tanaka-Ueguchi, M., Itoh, H., Nishimura, A., Swapan, D.,
- Tomita, M., Ishii, K. (2017) Genetisk ytelse av semidwarfing allelen SD1 avledet fra en japansk riskultivar og minimumskrav til å detektere sin enkeltnukleotidpolymorfisme med miSeq hele-genom Ssequencing. BioMed Research International.