Dele:



Hva er replikasjonsgaffelen?
den replikasjonsgaffel Det er punktet der DNA-replikasjon oppstår, det kalles også et vekstpunkt. Den har en Y-form, og etter hvert som replikering fortsetter, blir hårnålen forskjøvet av DNA-molekylet.
DNA-replikasjon er den cellulære prosessen som involverer duplisering av genetisk materiale i cellen. Strukturen av DNA er en dobbel helix, og for å replikere innholdet må den åpnes. Hver av strengene vil være en del av den nye DNA-kjeden, siden replikasjon er en semikonservativ prosess.
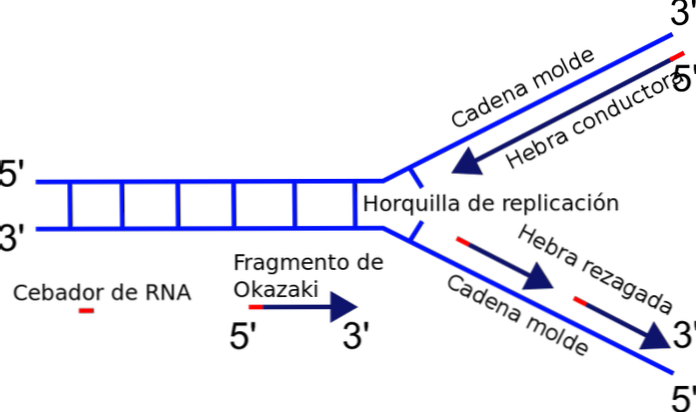
Replikasjonsgaffelen er dannet nøyaktig mellom overgangen mellom malen eller formkjeder separat og nylig dupleks-DNA som ikke er blitt duplisert. Ved initiering av DNA-replikasjon kan en av strengene enkelt dupliseres, mens den andre strengen står overfor et polaritetsproblem.
Enzymet som er ansvarlig for polymerisering av kjeden - DNA-polymerasen - syntetiserer bare DNA-strengen i 5'-3'-retningen. Således er en streng kontinuerlig og den andre lider en diskontinuerlig replikasjon, som genererer fragmenter av Okazaki.
index
- 1 Replikasjon av DNA og replikasjonsgaffel
- 1.1 Enveis og toveisreplikasjon
- 1.2 Enzymer involvert
- 1.3 Start av replikasjon og gaffelformasjon
- 1.4 Forlengelse og bevegelse av gaffelen
- 1.5 Oppsigelse
- 2 DNA-replikasjon er semikonservativ
- 3 Polaritetsproblemet
- 3.1 Hvordan polymerasen fungerer?
- 3.2 Produksjon av Okazaki-fragmenter
- 4 referanser
Replikasjon av DNA og replikasjonsgaffel
DNA er molekylet som inneholder den nødvendige genetiske informasjonen for alle levende organismer - med unntak av noen virus.
Denne store polymer bestående av fire forskjellige nukleotider (A, T, G og C) ligger i kjernen av eukaryotisk, i hver av cellene som utgjør vevet i disse vesener (bortsett fra modne røde blodceller fra pattedyr, mangler av kjernen).
Hver gang en celle deler seg, må DNA'et replikeres for å kunne opprette en dattercell med genetisk materiale.
Enveis og toveisreplikasjon
Replikasjon kan være ensrettet eller toveis, avhengig av dannelsen av replikasjonsgaffelen ved opprinnelsesstedet.
Logisk, når det gjelder replikasjon i en retning, blir det bare en gaffel dannet, mens toveisformene i toveisreplikasjon dannes.
Enzymer involvert
For denne prosessen er det nødvendig med et komplekst enzymatisk maskineri som fungerer raskt, og som kan replikere DNA på en presis måte. De viktigste enzymene er DNA-polymerase, DNA-primase, DNA-helikase, DNA-ligase og topoisomerase.
Start av replikering og formasjon av gaffelen
DNA-replikasjon starter ikke på noe tilfeldig sted i molekylet. Det er spesifikke regioner i DNA som markerer begynnelsen på replikasjonen.
I de fleste bakterier har bakteriekromosomet et enkelt AT-rikt utgangspunkt. Denne sammensetningen er logisk, siden den letter åpningen av regionen (AT-parene er forbundet med to hydrogenbroer, mens GC-paret med tre).
Når DNA begynner å åpne, dannes en Y-formet struktur: replikasjonsgaffelen.
Forlengelse og bevegelse av gaffelen
DNA-polymerase kan ikke starte syntesen av datterkjeder fra grunnen. Du trenger et molekyl som har en 3'-ende slik at polymerasen har hvor du skal begynne å polymerisere.
Denne gratis 3'-enden tilbys av et lite molekyl av nukleotider som kalles primer eller primer. Den første virker som en slags krok for polymerasen.
Med replikasjonsforløpet har replikasjonsgaffelen evnen til å bevege seg langs DNA-en. Plasseringen av replikasjonsgaffelen forlater to enkeltbånds DNA-molekyler som danner dannelsen av dobbeltbånds-dattermolekylene.
Gaffelen kan gå videre takket være virkningen av helikaseenzymer som uncoilerer DNA-molekylet. Dette enzymet bryter hydrogenbindingene mellom baseparene og tillater forskyvning av gaffelen.
ferdigstillelse
Replikasjonen avsluttes når de to gaflene er ved 180 ° C fra opprinnelsen.
I dette tilfellet snakker vi om som strømmer i prosessen for replikasjon i bakterier, og det bør bemerkes at prosessen med å vri den sirkulære molekyl som omfatter replikasjon. Topoisomeraser spiller en viktig rolle i avviklingen av molekylet.
DNA-replikasjon er semikonservativ
Har du noen gang lurt på hvordan replikasjon skjer i DNA? Det vil si at en annen dobbelthelix må oppstå fra den dobbelte helixen, men hvordan skjer det? I flere år var dette et åpent spørsmål blant biologer. Det kan være flere permutasjoner: to gamle tråder sammen og to nye sammen, eller en ny tråd og en gammel for å danne dobbeltspiralen.
I 1957 ble dette spørsmålet besvart av forskere Matthew Meselson og Franklin Stahl. Replikasjonsmodellen foreslått av forfatterne var semikonservativ.
Meselson og Stahl uttalte at resultatet av replikasjon er to dobbeltstrengede DNA-molekyler. Hver av de resulterende molekylene er sammensatt av en gammel streng (fra mor eller innledende molekyl) og en nylig syntetisert ny streng..
Polaritetsproblemet
Hvordan polymerase fungerer?
DNA-helixen er dannet av to kjeder som går på en antiparallell måte: Man går i 5'-3'-retningen og en annen 3'-5 '.
Den mest fremtredende enzym replikasjonsprosessen er DNA-polymerase, som er ansvarlig for å katalysere bindingen av nye nukleotider vil bli lagt til kjeden. DNA-polymerasen kan bare strekke kjeden i 5'-3'-retningen. Dette faktum hindrer samtidig duplisering av kjedene i replikasjonsgaffelen.
Hvorfor? Tilsetningen av nukleotider skjer ved den frie enden 3 'der en hydroksylgruppe (-OH) er funnet. Således kan bare en av kjedene lett bli forsterket ved terminalt tillegg av nukleotidet til 3'-enden. Dette kalles ledende eller kontinuerlig streng.
Produksjon av Okazaki-fragmenter
Den andre tråden kan ikke forlenges, fordi den frie enden er det 5 'og ikke den 3' og en hvilken som helst polymerase katalyserer tilsetning av nukleotider til 5 'ende. Problemet løses med syntesen av flere korte fragmenter (nukleotider 130 til 200), hver i retningen normal replikering 5 'til 3'.
Denne diskontinuerlige syntesen av fragmenter ender med foreningen av hver av delene, en reaksjon katalysert av DNA-ligase. Til ære for oppdageren av denne mekanismen, Reiji Okazaki, blir de små segmentene syntetisert kalt fragmenter av Okazaki.
referanser
- Alberts, B., Bray, D., Hopkin, K., Johnson, A. D., Lewis, J., Raff, M., ... & Walter, P. (2015). Nødvendig cellebiologi. Garland Science.
- Cann, I. K., & Ishino, Y. (1999). Archaeal DNA replikasjon: identifisere stykkene for å løse et puslespill. genetikk, 152(4), 1249-67.
- Cooper, G. M., og Hausman, R. E. (2004). Cellen: Molekylær tilnærming. Medicinska naklada.
- Garcia-Diaz, M., & Bebenek, K. (2007). Flere funksjoner av DNA-polymeraser. Kritiske vurderinger i plantevitenskap, 26(2), 105-122.
- Lewin, B. (2008). gener IX. Mc Graw-Hill Interamericana.
- Shcherbakova, P.V., Bebenek, K., & Kunkel, T. A. (2003). Funksjoner av eukaryotiske DNA-polymeraser. Vitenskapens SAGE KE, 2003(8), 3.
- Steitz, T. A. (1999). DNA-polymeraser: strukturell mangfold og felles mekanismer. Journal of Biological Chemistry, 274(25), 17395-17398.
- Watson, J. D. (2006). Molekylærbiologi av genet. Ed. Panamericana Medical.
- Wu, S., Beard, W. A., Pedersen, L.G., og Wilson, S.H. (2013). Strukturell sammenligning av DNA-polymerasarkitektur antyder en nukleotidgateway til polymeraseaktivt sted. Kjemiske vurderinger, 114(5), 2759-74.