Dele:



Cerex Cortex-lag, celler og funksjoner (med bilder)
den hjernebarken eller cerebral cortex er det nervøse vevet som dekker overflaten av hjernehalvene. Sa andre form, den utgjør den mest overlegne delen av hjernen.
Denne hjernestrukturen når sin maksimale utvikling i primater, er mindre utviklet i andre dyr og er relatert til utviklingen av kognitive og intellektuelle aktiviteter mer komplekse.
Den cerebrale cortex er et grunnleggende hjerneområde for menneskers funksjon. I denne regionen utføres funksjoner som oppfatning, fantasi, tanke, dom eller avgjørelse.
Anatomisk består den av en serie tynne lag bestående av grå materiale, som ligger over en bred samling av hvite sager.
Den cerebrale cortex vedtar en konvolutt form, så hvis den skulle strekke seg, ville den ha en meget omfattende masse. Nærmere bestemt tyder forskningen på at totalarealet av hjernebarken kan bestå av ca. 2500 kvadratkimeter.
På samme måte er denne store massen av hjerne preget av å inneholde et stort antall neuroner i sitt indre. På en generell måte er det postulert at i hjernebarken at det er om lag 10 milliarder nevroner, noe som ville gi omtrent 50 billioner synapser.
Hovedtrekkene til hjernebarken er forklart nedenfor. Dens lag, dets nevroner og dets funksjonelle organisasjon er spesifisert, og funksjonene som utføres i denne regionen av hjernen, blir vurdert.
index
- 1 Kjennetegn på hjernebarken
- 2 lag
- 2.1 Molekylært lag
- 2.2 Eksternt granulært lag
- 2.3 Eksternt pyramide lag
- 2.4 Internt granulært lag
- 2.5 lymfeknutelag
- 2.6 Flerskiktslag
- 3 Funksjonell organisering
- 3.1 Følsomme områder
- 3.2 Motorområder
- 3.3 Områder
- 4 nerveceller
- 4,1 pyramidale celler
- 4.2 Stellatceller
- 4.3 Spindelceller
- 4.4 Cajal horisontale celler
- 4,5 Martinotti celler
- 5 referanser
Kjennetegn på hjernebarken
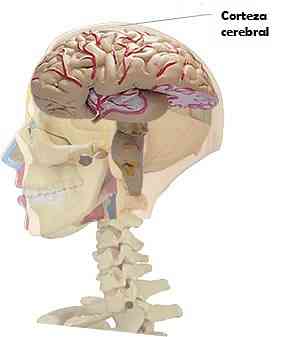
Den cerebrale cortex av pattedyr er representert av et ark med grå materiale, som dekker de to hjernehalvene.
Den består av en svært kompleks struktur hvor forskjellige sensoriske organer er representert i bestemte områder eller områder, som kalles primære sensoriske områder.
Hver av de fem sansene som mennesker har (syn, berøring, lukt, smak og berøring) utvikler seg i en spesifikk del av cortex. Det vil si at hver sensorisk modalitet har et avgrenset territorium i hjernebarken.
Bortsett fra de sensoriske områdene har hjernebarken også flere sekundære somatiske, forening og motorregioner. På disse områdene utvikles kortikale og assosiasjonssamfunnssystemer, noe som gir opphav til læring, minne og atferd.
I denne forstand betraktes hjernebarken som et spesielt relevant område når man utvikler de overordnede aktivitetene i den menneskelige hjerne.
De mest avanserte og forseggjorte menneskelige prosesser som resonnement, planlegging, organisering eller forening utføres i forskjellige områder av hjernebarken.
Av denne grunn utgjør hjernebarken en struktur som oppnår maksimal kompleksitet fra et menneskelig perspektiv. Den cerebrale cortex er resultatet av en sakte evolusjonær prosess som kunne ha startet mer enn 150 millioner år siden.
lagene
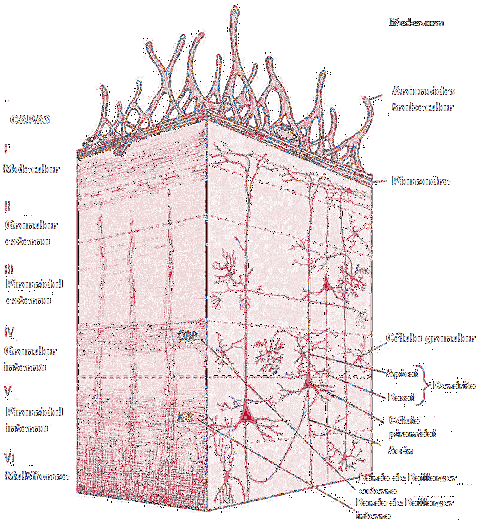
Hovedkarakteristikken til hjernebarken er at den utgjøres av forskjellige lag av grått materiale. Disse lagene utgjør strukturen i skorpen og definerer dens strukturelle og funksjonelle organisasjon.
I tillegg er lagene i hjernebarken karakterisert ved at de ikke bare er definert fra et strukturelt synspunkt, men også fra et fylogenetisk perspektiv.
Det vil si at hvert lag i hjernebarken tilsvarer et annet evolusjonært øyeblikk. Ved begynnelsen av menneskelige arter var hjernen mindre utviklet og cortexen hadde færre lag.
Gjennom utviklingen av arten har disse lagene økt, faktum som er relatert til økningen av kognitive og intellektuelle evner hos mennesker over tid.
Molekylært lag
Det molekylære laget, også kjent som plexiformlaget, er den mest overfladiske regionen i hjernebarken, og derfor er den nyeste utbruddet.
Den består av et tett nettverk av nervefibre som er tangentielt orientert. Disse fibrene er avledet fra dendriter av pyramidale og fusiformceller, akselene til stellatet og Martinotti-cellene.
Afferente fibre som kommer fra thalamus, forening og kommisjoner kan også bli funnet i molekylærlaget. Å være den mest overfladiske regionen i cortexen, i molekylærlaget etableres et stort antall synapser mellom forskjellige neuroner.
Eksternt granulært lag
Det ytre granulære lag er den nest mest overfladiske regionen i cortexen og ligger under molekylærlaget. Den inneholder et stort antall små pyramide- og stellatceller.
Dendritene til de ytre granulære lagcellene ende opp i molekylærlaget og axonene går inn i dypere lag av hjernebarken. Av den grunn er det ytre granulære lag sammenkoplet med de forskjellige områder av cortex.
Eksternt pyramide lag
Det ytre pyramidale laget, som navnet antyder, består av pyramidale celler. Den er preget av en uregelmessig form, det vil si at lagets størrelse øker fra overflaten til den dypeste grensen.
Dendrittene til neuronene i pyramidlaget går til molekylærlaget og axonene beveger seg som fremspring, forening eller kommissurale fibre til det hvite stoffet som ligger mellom lagene i hjernebarken.
Internt granulært lag
Det indre granulære laget består av stellatceller som er arrangert i en svært kompakt form. Den har en høy konsentrasjon av horisontalt anordnede fibre kjent som Baillarger ytre bånd.
Ganglion lag
Ganglionlaget eller det indre pyramide lag inneholder svært store og mellomstore pyramidale celler. På samme måte inneholder de et høyt antall fibrene arrangert horisontalt som danner det indre Baillarger-bandet.
Multiformellag
Endelig inneholder det flerlagige laget, også kjent som det polymorfe cellelaget, fusiformceller. På samme måte inneholder den modifiserte pyramidale celler som inneholder en trekantet eller ovoid cellelegeme.
Mange av nervefibrene i flerskiktslaget går inn i det underliggende hvite saken og forbinder laget med mellomliggende områder.
Funksjonell organisering
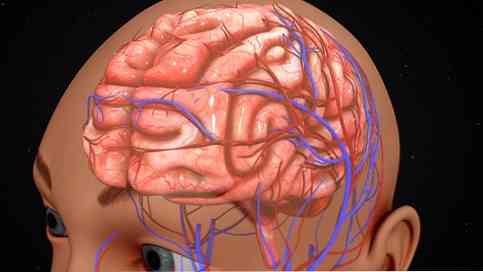
Den cerebrale cortex kan også organiseres i henhold til aktivitetene som utføres i hver region. I denne forstand bestemte områder av hjernebarksprosessen spesifikke signaler av en følsom, motorisk og foreningskarakter..
Følsomme områder
Sanseområder er regioner i hjernebarken som mottar informasjon av følsom karakter og er nært knyttet til oppfatningen.
Informasjonen åpner hjernebarken hovedsakelig gjennom den bakre halvdel av begge hjernehalvene. De primære områdene inneholder de mest direkte forbindelser med perifere sensoriske reseptorer.
På den annen side er sekundære sensoriske og forening områder vanligvis i nærheten av de primære områdene. Generelt mottar de informasjon fra både de primære tilknytningsområdene og de nedre områdene i hjernen..
Hovedoppgaven til områdene for tilknytning og sekundære områder er å integrere sensitive opplevelser for å generere mønstre av anerkjennelse og oppførsel. De viktigste følsomme områdene i hjernebarken er:
- Det primære somatosensoriske området (område 1, 2 og 3).
- Det primære visuelle området (område 17).
- Det primære lydområde (område 41 og 42).
- Det primære smaksområdet (område 43).
- Det primære olfaktoriske området (område 28).
Motorområder
Motorområdene er i den fremre delen av halvkulen. De er ansvarlige for å igangsette hjerneprosesser knyttet til bevegelse og gi opphav til slike aktiviteter.
De viktigste motorområdene er:
- Det primære motorområdet (område 4).
- Området for borspråk (område 44 og 45).
Områder
Områdene av hjernebarken korrelerer med de mer komplekse integrasjonsfunksjonene. Disse områdene utfører aktiviteter som minnes- og kognitionsprosesser, styring av følelser og utvikling av resonnement, vilje eller dom.
På samme måte spiller foreningens områder en spesielt viktig rolle i utviklingen av personlighet og karaktertrekk hos mennesker. På samme måte er det en hjernegruppe avgjørende for bestemmelsen av intelligens.
Områdene inkluderer bestemte motorområder, samt spesifikke sensoriske områder.
Nerveceller

Den cerebrale cortex har et stort utvalg av celler inne i den. Spesifikt har fem forskjellige typer nevroner blitt spesifisert i denne regionen av hjernen.
Pyramidale celler
Pyramidceller er nevroner som er preget av å ha en pyramideform. De fleste av disse cellene inneholder en diameter mellom 10 og 50 mikrometer.
Det er imidlertid også store pyramidale celler. Disse er kjent som Betz-celler og kan ha en diameter på opptil 120 mikrometer.
Både små pyramidceller og store pyramidceller finnes i motorisk precentral sirkonløsning og utfører hovedsakelig aktiviteter relatert til bevegelse.
Stellate celler
Stellatceller, også kjent som granulosa-celler, er små nevroner. De har vanligvis en diameter på ca 8 mikrometer og har en polygonal form.
Spindelceller
Fusiformceller er nevroner som har sin vertikale langsgående akse på overflaten. De er konsentrert hovedsakelig i de dypere kortikale lagene i hjernen.
Axonene fra disse nevronene stammer fra underkroppens nedre del og er rettet mot den hvite saken som fremspring, forening eller kommissorisk fiber..
Cajal horisontale celler
De horisontale cellene av cajal er små fusiformceller som er orientert horisontalt. De er i de mest overfladiske lag av hjernebarken og spiller en avgjørende rolle i utviklingen av denne hjerneregionen.
Slike nevronene ble oppdaget og beskrevet av Ramon y Cajal slutten av nittende århundre, og senere undersøkelser viste som cellene er viktig for å koordinere neuronal aktivitet.
For å oppnå sin posisjon i hjernebarken må de horisontale cellene av kjeve migrere på en koordinert måte under embryogenesen av hjernen. Det vil si at disse nevronene reiser fra deres fødested til overflaten av hjernebarken.
Når det gjelder molekyl mønster av disse neuronene, Victor Borrell og Oscar Marin Neuroscience Institute of Alicante, viste at de horisontale celler av cajal oppviser en orientering av neuronale lag av hjernebarken under embryonal utvikling.
Faktisk kommer dispersjonen av disse cellene opp i løpet av de første faser av embryonisk utvikling. Cellene er født i forskjellige områder av hjernen og migrerer til hjernens overflate for å dekke det helt.
Endelig har det nylig blitt vist at meningeal membraner har andre funksjoner enn den beskyttende antatt at i utgangspunktet. Hjernehinnene tjene som substrater eller horisontale bane Cajal celler for tangential migrering av overflaten av cortex.
Martinotti celler
De siste nevronene som utgjør den cerebrale cortex nevronaktivitet, er de velkjente Martinotti-cellene. De består av små multiforme nevroner tilstede på alle nivåer i hjernebarken.
Disse nevronene skylder deres navn til Carlo Martinotti, en studentforsker av Camilo Golgi som oppdaget eksistensen av disse cellene i hjernebarken.
Martinotti-celler kjennetegnes ved å være multipolære neuroner med korte arboreserende dendriter. De blir spredt gjennom flere lag av hjernebarken og sender sine axoner til molekylærlaget, hvor axonale arboriseringer dannes.
Nylig forskning på disse nevronene har vist at Martinotti-celler deltar i hjernens hemmeringsmekanisme.
Nærmere bestemt, når en pyramideformet neuron (som er den vanligste typen av nevroner i den cerebrale cortex) starter overexcited, Martinotti cellene begynner å overføre inhiberende signaler til nervecellene rundt.
I denne forstand, følger det at epilepsi kan være sterkt forbundet med et underskudd av Martinotti celler eller et underskudd på aktiviteten til disse neuronene. I disse øyeblikkene er den nervøse overføringen av hjernen ikke lenger regulert av disse cellene, noe som forårsaker ubalanse i cortexens funksjon..
referanser
- Abeles M, Goldstein MH. Funksjonell arkitektur i katt primær auditiv cortex. Columnar organisasjon og organisasjon i henhold til dybden. J Neurophysiol 1970; 33: 172-87.
- Blasdel GG, Lund JS. Oppsigelse av afferente axoner i makaque striat cortex. J Neurosci 1983; 3: 1389-413.
- Chang HT. Kortikale nevroner med særlig referanse til apikale dendriter. Cold Spring Harb Symp Quant Biol 1952; 17: 189-202.
- Fra Felipe J. Lysekrone celler og epilepsi. Brain 1999; 122: 1807-22.
- Ramón og Cajal S. Neue Darstellung fra histologischen Bau des Centralnerevensystem. Arch Anat Physiol 1893: 319-428.
- Rubenstein JLR, Rakic P. Genetisk kontroll av kortikal utvikling. Cereb Cortex 1999; 9: 521-3.