Dele:



Felles celletyper og deres egenskaper
den celleforbindelser de er kontaktbroer som eksisterer mellom cytoplasmatiske membraner mellom tilstøtende celler eller mellom en celle og matrisen. Leddene avhenger av hvilken type vev som studeres, og markerer forbindelsene mellom epitel-, muskel- og nervceller.
I cellene er det molekyler knyttet til adhesjonen mellom dem. Imidlertid er det nødvendig med ytterligere elementer som øker stabiliteten av bindingen av vev. Dette oppnås med cellulære veikryss.
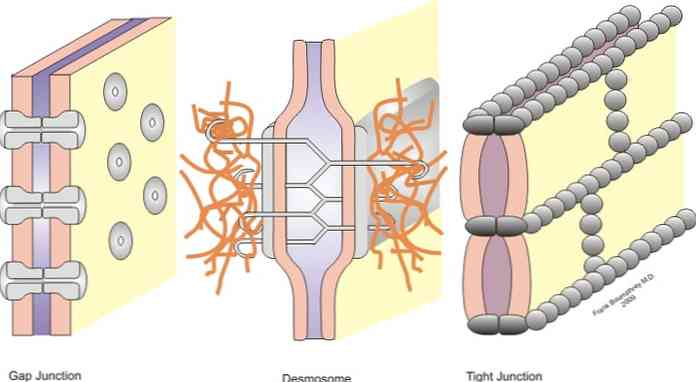
Leddene er klassifisert i symmetriske ledd (smale ledd, desmosomer i beltet og spaltene) og asymmetriske ledd (hemidesmosomer).
Smale veikryss, belte desmosomer, punkt desmosomer og hemidesmosomer er ledd som tillater forankring; mens kryssene i spalten oppfører seg som broer av forening mellom nabo-cellene, slik at utveksling av oppløsninger mellom cytoplasmaene.
Bevegelsen av løsemidler, vann og ioner skjer gjennom og mellom individuelle cellulære komponenter. Dermed er det den transcellulære vei som styres av en rekke ledninger og transportører. I motsetning til den paracellulære banen, som reguleres av kontakter mellom celler - det vil si celleforbindelser.
I planter finner vi cellulære veikryss som ligner slitekryss, kalt plasmododer. Selv om de er forskjellige i strukturen, er funksjonen den samme.
Fra medisinsk synspunkt fører visse mangler i celleforbindelser til oppkjøpte eller arvelige sykdommer forårsaket av skade på epitelbarrieren.
index
- 1 Egenskaper
- 2 typer
- 2.1 - Smarte tilkoblinger
- 2.2 - Spalt eller mellomrom
- 2.3 - Forankring eller vedhengende tilkoblinger
- 2.4 -Hemidesmosomer
- 3 Cellulære veikryss i planter
- 4 medisinsk perspektiv
- 5 referanser
funksjoner
Levende organismer består av diskrete og varierte strukturer kalt celler. Disse er avgrenset av en plasmamembran som holder dem skilt fra det ekstracellulære miljøet.
Men selv om de er bestanddelene til levende vesener, ser de ikke ut som murstein, da de ikke er isolert fra hverandre.
Cellene er elementer som kommuniseres med hverandre og med det ekstracellulære miljøet. Derfor må det være en måte for celler å danne vev og kommunisere, mens membranen forblir intakt.
Dette problemet er løst takket være forekomsten av celleforbindelser som eksisterer i epitelene. Disse veikryssene dannes mellom to tilstøtende celler klassifiseres i henhold til funksjonen til hver i symmetriske og asymmetriske ledd.
Asymmetriske tilkoblinger tilhører hemidesmosomer, og ved symmetriske tilkoblinger lukker forbindelser, et desmosom på et belte, desmosomer og slitforbindelser. Neste vil vi beskrive i detalj hver av fagforeningene.
typen
-Smale veikryss

Smale veikryss, også kjent i litteraturen som okklusive veikryss, er sektorer i cellemembranene i nabokjeller som er nært forbundet - som navnet "smalt veikryss" indikerer.
Under gjennomsnittlige forhold separeres cellene med en avstand på 10 til 20 nm. Imidlertid, når det gjelder smale veikryss, blir denne avstanden betydelig redusert og membranene til begge cellene berøres eller til og med slått sammen.
Et typisk smalt veikryss befinner seg mellom sideveggene til nabo-cellene i en minste avstand fra deres apikale overflater.
I epitelvevet etablerer alle cellene fagforeninger av denne typen for å forbli forenet. I denne interaksjonen ligger cellene som danner et mønster som minner om en ring. Disse fagforeningene dekker hele omkretsen.
Proteiner involvert i stramme kryss
Ocludina og Claudina
Områdene av nær kontakt omgir hele overflaten av cellen. Disse områdene danner anastomoserende kontaktstrimler av transmembranproteinene kjent som occludin og claudin. Begrepet anastomose refererer til foreningen av visse anatomiske elementer.
Disse to proteinene tilhører gruppen tetraespaniner. De er preget av å ha fire transmembrane domener, to eksterne sløyfer og to relativt korte cytoplasmatiske haler.
Det har blitt observert at okkludin interagerer med fire andre proteinmolekyler, kalt occludin av zonulaen og forkortet som ZO. Denne siste gruppen inkluderer proteiner ZO 1, ZO 2, ZO 3 og afuna.
Claudin, derimot, er en familie av 16 proteiner som utgjør en serie lineære fibriller i de smale kryssene, noe som gjør at denne unionen kan ta rollen som en "barriere" i paracellulær banen..
Nectinas og JAM
Nektinene og adhesjonsmolekylene til fagforeningene (forkortet av akronym i engelsk JAM), vises også i de smale kryssene. Disse to molekylene finnes som homodimerer i det intracellulære rommet.
Nektinene er koblet til actinfilamenterne ved hjelp av afadinproteinet. Sistnevnte ser ut til å være avgjørende, da de i slettingen av genet som koder for afadin hos gnagere, fører til embryoens død.
Funksjoner av smale veikryss
Disse typer celle-til-celle krysser utfører to viktige funksjoner. Den første er å bestemme polariteten til cellene i epitelet, separere det apikale domenet fra basolateralt og forhindre en unødig diffusjon av lipider, proteiner og andre biomolekyler..
Som vi nevnte i definisjonen, grupperes cellene i epitelet i en ring. Denne strukturen skiller den apikale overflaten av cellen fra de laterale og basale, som etablerer differensieringen mellom domenene.
Denne separasjonen anses som en av de viktigste begrepene i studiet av epitelia fysiologi.
For det andre hindrer de stramme kryssene fri passasje av stoffer gjennom laget av epitelceller, noe som overskrider en barriere mot den paracellulære banen..
-Spalt eller gapskryss

Spalt- eller mellomromskryssene er funnet i områder som ikke er begrenset til cytoplasmatisk membran mellom nabokeller. I en delt knutepunkt kobles cytoplasmaene til cellene og det oppstår en fysisk forbindelse hvor små molekyler kan forekomme.
Denne klassen av ledd finnes i nesten alle epitelene, og i andre typer vev, hvor de tjener ganske varierte formål..
For eksempel kan i flere vev slitekryssene åpne eller lukke som svar på ekstracellulære signaler, slik det er tilfelle med nevrotransmitterdopaminen. Tilstedeværelsen av dette molekylet reduserer kommunikasjonen mellom en klassenuron i retina, som respons på den økte intensiteten av lys.
Proteiner involvert i spaltning ledd
Spaltningspunkter er dannet av proteiner kalt connexiner. Dermed blir en "koneksjon" oppnådd ved forening av seks konnexin-monomer. Denne strukturen er en hul sylinder som passerer gjennom den cytoplasmiske membranen.
Forbindelsene er anordnet slik at en rørledning oppstår mellom cytoplasmaene til de tilstøtende celler. I tillegg har konnexons en tendens til å aggregere og danne en slags plater.
Funksjoner av slit ledd
Takket være dannelsen av disse bindingene kan det forekomme bevegelse av visse molekyler mellom nabolagene. Størrelsen på molekylet som skal transporteres er avgjørende, den optimale diameter er 1,2, slik som kalsiumioner og cyklisk adenosinmonofosfat.
Spesielt er det de uorganiske ionene og de vannløselige molekylene som kan overføres fra en cellulær cytoplasma til den sammenhengende cytoplasma.
Kalsiumkonsentrasjoner spiller en avgjørende rolle i denne kanalen. Når kalsiumkonsentrasjonen øker, har de aksiale kanalene en tendens til å lukke.
På denne måten deles slitsleddene aktivt i prosessen med elektrisk og kjemisk kobling mellom celler, som forekommer i hjertemuskelcellene, som er ansvarlige for overføring av elektriske impulser.
-Forankring eller vedhengende tilkoblinger

Under de smale leddene finner vi forankringsleddene. Vanligvis ligger disse i nærheten av epitelens apikale overflate. I denne gruppen kan vi skille tre hovedgrupper, zonula adherens eller desmosoma i beltet, makula adherens eller desmosome punkt og desmosome.
I denne typen krysser er tilstøtende cellemembraner som er forbundet med zonules og adherens macules separert av en cellulær avstand som er relativt bred - hvis vi sammenligner dem med minimumsrom som eksisterer ved smale kryss..
Det intercellulære rom er okkupert av proteiner som tilhører familien av cadheriner, desmogleiner og desmocolinas knyttet til cytoplasmatiske plater som har andre proteiner kalt desmoplaquina, placoglobina og placofilina.
Klassifisering av forankringsledd
Zonula adherens
Som i tilfelle av smale ledd, i forankringsleddene, observerer vi også arrangementsmønsteret i form av en ring eller et belte. Zonula-adherens er forbundet med actinmikrofilm, gjennom samspillet mellom to proteiner: kadheriner og kateniner.
Macula adherens
I noen tilfeller er denne strukturen bare kjent som desmosoma, det er en punkteringskryss som er forbundet med mellomliggende filamenter dannet av keratin. I denne sammenheng kalles nevnte keratinstrukturer "tonofilimanetos". Filamentene strekker seg fra ett punkt til et annet i epitelceller.
Punkt desmosomer
Disse gir styrke og stivhet til epitelceller. Dermed antas det at dets hovedfunksjon er relatert til styrking og stabilisering av tilstøtende celler.
Desmosomer kan sammenlignes med en slags nitter eller sveiser, siden de ligner separate små flekker og ikke kontinuerlige band.
Vi finner denne type skjøter innføyet disker som binder kardiomyocytter i hjertemuskelen og i hjernehinnene lining den ytre overflaten av hjernen og ryggmargen.
-hemidesmosome

Hemidesmosomer faller inn i kategorien av asymmetriske veikryss. Disse strukturene har funksjonen til å forankre det basale domenet til epitelcellen med den underliggende basale lamina.
Begrepet hemidesmosom brukes fordi denne strukturen virker, bokstavelig talt, "medium" desmosoma. Men med utgangspunkt i deres biokjemiske sammensetning er begge fagforeninger helt forskjellige.
Det er viktig å klargjøre at desmosomene er ansvarlige for å feste en nærliggende celle med en annen, mens funksjonen til hemidesmosomet er å bli med i cellen med den basale lamina.
I motsetning til adherens makula eller desmosom, hemidesmosomes har en annen struktur, som består av: et cytoplasmatisk ark forbundet med intermediære filamenter og plate ytre membraner, som er ansvarlig for å feste hemidesmosome med basal lamina, gjennom en ankerfilament.
En funksjon er hemidesmosomes øke den totale stabiliteten av epitelvev, takket være tilstedeværelsen av cytoskjelett intermediære filamenter festet til komponenter av basal lamina.
Cellforbindelser i planter
Plantedrykket mangler de fleste av cellekryssene som er beskrevet ovenfor, med unntak av en funksjonell motstykke som ligner slitekryss.
I planter er cytoplasmaene til tilstøtende celler forbundet med baner eller kanaler som kalles plasmodioder.
Denne strukturen skaper et kontinuum fra en plantecelle til den neste. Selv om det avviker strukturelt fra spaltkryssene, har de svært like roller, slik at passasje av små ioner og molekyler.
Medisinsk perspektiv
Fra medisinsk synspunkt er celleforeninger et relevant problem. Det har blitt funnet at mutasjoner i gener som koder for proteiner involvert i kryssene, blir oversatt til kliniske patologier.
For eksempel, hvis det er noen mutasjon i genet som koder for en bestemt type claudin (ett av proteinene som medierer interaksjons tilslutninger smal type) bevirker en sjelden sykdom hos menneske.
Dette er syndromet av tap av nyre magnesium, og symptomene inkluderer lav magnesium og anfall.
I tillegg har det blitt funnet at en mutasjon i genet som koder for protein nektin 1, er ansvarlig for cleft palatesyndrom eller spaltleppe. Denne tilstanden regnes som en av de vanligste misdannelsene hos nyfødte.
Mutasjoner i genet nectin en har også vært forbundet med annen tilstand som kalles ektodermal dysplasi påvirker hud, hår, negler og tenner til mennesker.
Pemphigus foliaceus er bulløs hudpatologien bestemt av autoantistoffer mot desmoglein 1, sentralt element som er ansvarlig for å opprettholde sammenhengen i epidermis.
referanser
- Alberts, B., Bray, D., Hopkin, K., Johnson, A. D., Lewis, J., Raff, M., ... & Walter, P. (2015). Nødvendig cellebiologi. Garland Science.
- Cooper, G. M., og Hausman, R. E. (2000). Cellen: Molekylær tilnærming. Sinauer Associates.
- Curtis, H., & Barnes, N. S. (1994). Invitasjon til biologi. Macmillan.
- Hill, R.W., Wyse, G. A., Anderson, M., Anderson, M. (2004). Dyrfysiologi. Sinauer Associates.
- Karp, G. (2009). Cell- og molekylærbiologi: konsepter og eksperimenter. John Wiley & Sons.
- Kierszenbaum, A., & Tres, L. (2016). Histologi og cellebiologi: en introduksjon til patologi. Elsevier Brasil.
- Lodish, H., Berk, A., Darnell, J. E., Kaiser, C.A., Krieger, M., Scott, M.P., ... & Matsudaira, P. (2008). Molekylærcellebiologi. Macmillan.
- Voet, D., & Voet, J. G. (2006). biokjemi. Ed. Panamericana Medical.